

Maternal Immune Activation During Pregnancy Occurs Hundreds to Tens of Thousands of Times Under CDC’s Current Recommended Schedule
Known immunogenic epitopes number in the hundreds. Jake Scott's "few antigens" position is naive and misleading.

Maternal immune activation (MIA) is no longer a speculative construct. Across experimental immunology, placental biology, and human epidemiology, the signal is consistent: immune activation during pregnancy can alter fetal developmental trajectories without fetal infection. Cytokine signaling, acute-phase responses, and placental endocrine disruption form the mechanistic bridge (Smith, 2007; Hsiao, 2011; Wu, 2017).
This article integrates that evidence on the first pregnancy-specific, CDC-schedule-aligned quantitative analysis of vaccine antigen and epitope exposure explicitly framed in the context of maternal immune activation.
This key analysis corrects a persistent analytical error: counting pathogens as “antigens.” Proteins are antigens. Peptides are the operational units of immune recognition. When that biology is respected, the magnitude of immune activation opportunities during pregnancy becomes visible.
===================================================================
Clarifying analytical terms used throughout this article.
This analysis distinguishes three non-interchangeable quantities that are often conflated in public discussions of vaccine immunology.
Protein antigens refer to the distinct protein molecules delivered by a vaccine formulation.
Possible epitopes refer to the total number of linear peptide windows (e.g., 9-mer and 15-mer sequences) that exist within those proteins and define the upper bound of antigen-processing opportunity.
Likely epitopes refer to peptide sequences that have been experimentally demonstrated to be processed, presented, and recognized by human CD4 T cells under defined assay conditions.
These categories are analytically distinct. Conflating them obscures scale, mechanism, and biological relevance.
===================================================================
What follows proceeds in three steps. First, the evidence base that MIA constitutes a developmental risk. Second, the empirical reality that serious infections during pregnancy are uncommon in modern settings, making iatrogenic immune activation a relevant comparison class. Third, a rigorous antigen and epitope analysis of influenza, Tdap, RSV, and COVID‑19 vaccines as recommended during pregnancy, including why claims that these schedules involve “very few antigens” are biologically incorrect.
Maternal Immune Activation as a Developmental Risk
MIA refers to a maternal immune state characterized by cytokine release, innate immune activation, and downstream placental signaling that alters the fetal environment. It does not require fetal infection. In animal models, maternal exposure to immune triggers such as viral mimetics is sufficient to produce lasting changes in offspring behavior, cortical organization, synaptic development, and gene expression (Smith, 2007). Interleukin‑6 signaling has been shown to be both sufficient and necessary in canonical models, and placental JAK/STAT activation has been identified as a critical transducer that links maternal immune signaling to altered fetal growth factor and endocrine output (Hsiao, 2011). Placental IL‑6 signaling also modulates fetal brain development and later behavior (Wu, 2017).
Human data converge on the same conclusion at the level of association. Meta‑analyses and large population cohorts consistently show elevated risks of autism spectrum disorder, schizophrenia, psychosis-spectrum disorders, and other neurodevelopmental outcomes following maternal infection or fever during pregnancy (Tioleco, 2021; Antoun, 2021; Zhou, 2021; Saatci, 2021). Effect sizes are modest but robust, typically in the range of 1.2–1.6 depending on outcome, timing, and severity. Biomarker studies strengthen causal plausibility: elevated maternal C‑reactive protein during early gestation predicts later autism and schizophrenia diagnoses in offspring (Brown, 2014; Canetta, 2014). Timing matters. First‑trimester viral immune activation and second‑trimester bacterial activation produce stronger signals than undifferentiated exposure categories (Atladóttir, 2010), consistent with known windows of neurodevelopmental vulnerability.
These data establish a critical point for any risk analysis: immune activation during pregnancy is a biologically meaningful exposure class, not a null background condition.
Infection During Pregnancy Is Relatively Uncommon
In high‑income countries, severe infections requiring hospitalization during pregnancy are uncommon. Even outpatient infections with sustained fever represent a minority of pregnancies. This epidemiologic reality matters because it defines the baseline frequency of naturally occurring, clinically significant maternal immune activation.
By contrast, the CDC’s current recommendations explicitly encourage immune activation during pregnancy through vaccination against influenza, pertussis, tetanus, diphtheria, RSV, and COVID‑19. These recommendations are often justified by asserting that vaccines introduce “very few antigens,” especially when compared to natural infection. That assertion rests on a categorical error.
The Error: Counting Pathogens as Antigens
A recurring claim, articulated publicly by science communicators such as Jake Scott, is that vaccination during pregnancy introduces only a small number of antigens because each vaccine is treated as if it contained “one antigen,” often equated with a pathogen. This framing is biologically, and specifically, immunologically false.
The immune system does not recognize organisms. It recognizes molecular structures. Specifically, it recognizes protein- and peptide-derived epitopes presented on major histocompatibility complex molecules and, in the case of antibodies, conformational protein surfaces. A virus is not an antigen. A bacterium is not an antigen. Proteins are antigens, and peptides derived from those proteins are the operational units of T‑cell recognition.
Counting pathogens as antigens collapses molecular immunology into folk taxonomy. It erases the difference between a single purified protein and a multi‑protein toxoid vaccine. It is analytically equivalent to claiming that a book has “one word” because it has one cover.
Proteins, Peptides, and Epitope Opportunity
A protein of length L amino acids contains L–8 distinct overlapping 9‑mer peptide frames and L–14 overlapping 15‑mer frames. These windows define the upper bound of class I and class II T‑cell epitope opportunity space. Not all of these peptides will be processed, presented, or recognized. Immunology imposes strong filters. Antigen processing deletes most candidates. HLA binding deletes most of what remains. T‑cell receptor availability and immunodominance delete most of the rest.
But deleting is not the same as negating. What matters is scale. A protein 500–600 amino acids in length contains on the order of one thousand distinct linear peptide windows. Empirically, human epitope mapping studies repeatedly show that dozens of peptides per protein are presented and recognized across populations, and that individual donors typically mount responses to multiple distinct epitopes per antigen (Cassotta, 2020; Mateus, 2020; Parker, 2023).
Thus, the correct question is not whether “all epitopes are immunogenic.” They are not. The correct question is whether the antigenic and epitope load is biologically trivial. It is not.
Caveat: These epitope and peptide counts represent antigenic elements that are biologically capable of being generated, presented, and immunologically active in humans; maternal immune activation risk depends on this potential for immune engagement, not on whether every epitope is realized or dominant in every individual.
The CDC Pregnancy Vaccination Schedule: Antigen Reality
Under current CDC guidance, a pregnant woman may receive vaccination against influenza, Tdap, RSV, and COVID‑19 during a single pregnancy.
Seasonal influenza vaccines used in the United States are trivalent and contain three distinct hemagglutinin proteins. Each hemagglutinin is approximately 560–580 amino acids long, yielding roughly 1,100 distinct 9‑mer and 15‑mer peptide windows per protein. Influenza vaccination during pregnancy therefore introduces on the order of 3,300 peptide opportunities from hemagglutinin alone.
Tdap is the dominant antigenic contributor. Tetanus toxoid is a 1,315‑amino‑acid protein. Diphtheria toxoid is approximately 535 amino acids. Acellular pertussis vaccines include five pertussis toxin subunits plus additional proteins such as pertactin and filamentous hemagglutinin. FHA is particularly important: it is derived from a precursor exceeding 3,500 amino acids, with the secreted antigen corresponding to a large processed fragment. Depending on how one models the presented FHA sequence, it alone contributes between roughly 4,000 and over 7,000 peptide windows. Depending on whether BOOSTRIX or ADACEL is used, total Tdap protein antigen counts range from nine to eleven distinct proteins, with total 9‑mer plus 15‑mer peptide opportunity counts between approximately 11,600 and 15,300.
Maternal RSV vaccination with ABRYSVO introduces two stabilized prefusion F proteins, one from RSV‑A and one from RSV‑B. Each F protein is approximately 574 amino acids long, contributing roughly 1,126 peptide windows each, for a total of about 2,250 (van Bleek, 2004).
COVID‑19 mRNA vaccination introduces the SARS‑CoV‑2 spike protein, a 1,273‑amino‑acid antigen corresponding to roughly 2,500 distinct 9‑mer and 15‑mer peptide windows (Mateus, 2020; Parker, 2023).
When these components are summed, a single pregnancy following CDC recommendations is associated with approximately 15–17 distinct protein antigens and between roughly 19,800 and 23,400 distinct linear peptide opportunities. The range is driven almost entirely by reasonable uncertainty in how FHA processing is modeled.
This is not rhetoric. It is arithmetic applied to protein sequences.
Importantly, the experimentally enumerated “likely epitope” counts used later in this article represent lower-bound estimates, not comprehensive inventories. Individuals and families have “private” alleles - genetic variation that is not shared by many other people. These can change amino acid sequences and create family- or even individua-specific immunoepitopes that set them up for pathogenic priming due to exposures to pathogenic epitopes in vaccine or via infection. The total number known are also constrained by assay thresholds, cohort size, HLA diversity, and the absence of pregnancy-specific epitope-mapping studies. Absence of detection in these datasets does not imply absence of immunogenic potential.
(Article continues after image)…

Immunogenic Reality: Filtering Does Not Erase Scale
It is true, and important, that only a fraction of theoretical peptide windows become immunogenic. Human epitope mapping studies provide calibration.
All epitope-mapping data referenced here derive from non-pregnant human cohorts. Pregnancy substantially alters immune thresholds, regulatory T-cell balance, cytokine sensitivity, and placental immune surveillance. The direction and magnitude of these effects on antigen presentation and immunodominance are incompletely characterized, reinforcing the need for pregnancy-specific immune profiling rather than extrapolation alone.
For SARS‑CoV‑2 spike, human CD4 mapping studies identify on the order of 60–70 distinct epitopes across populations, while individual donors typically respond to approximately 10–30 epitopes (Mateus, 2020).
RSV F mapping studies identify roughly 30 immunodominant peptides (van Bleek, 2004). Tetanus toxoid studies show that fewer than 50 epitopes account for essentially all measurable CD4 responses, with approximately 28 accounting for 90 percent of the response (da Silva Antunes, 2018).
Influenza HA is similarly constrained: despite a long protein, CD4 memory concentrates on a small number of immunodominant peptide regions, as shown by both ex vivo and in vitro activation studies (Cassotta, 2020; Babon, 2012).
These data do not rescue the “few antigens” argument. They refute it. Even after all biological filters are applied, vaccines routinely generate double‑digit epitope recognition per antigen in individual humans and far larger repertoires across populations. Multiply that across 15–17 protein antigens administered during pregnancy and the immunologic burden is plainly nontrivial.
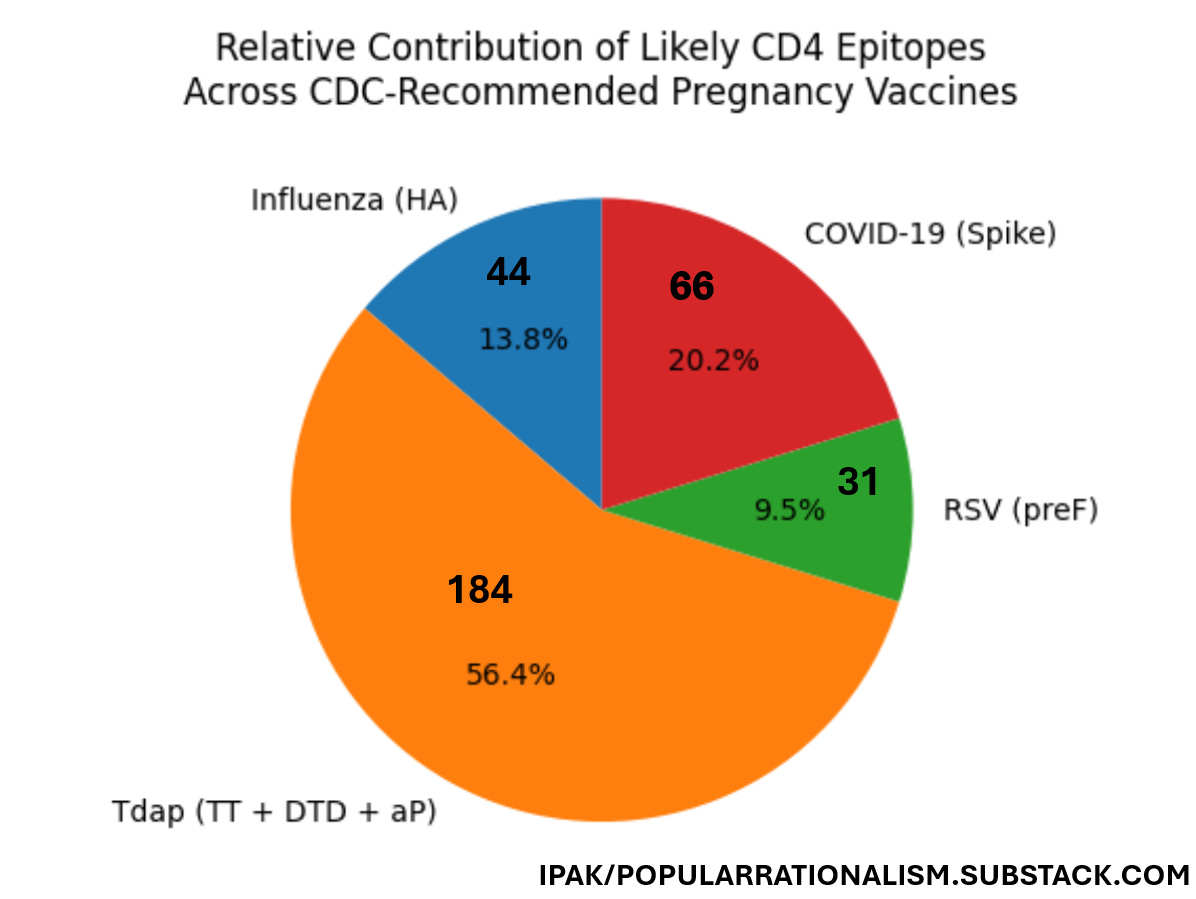
Figure 1. Conservative estimate of the number of CD4 epitopes per vaccine type. When experimentally enumerated human CD4 epitope data are considered, the distribution of likely epitope exposure is highly non-uniform across vaccines. Conservative literature-based counts indicate that the Tdap vaccine accounts for the majority of enumerated CD4 epitopes, followed by SARS-CoV-2 spike, influenza hemagglutinin, and RSV prefusion F. This distribution underscores that schedule composition—not merely the number of injections—determines immunologic burden.
From Antigen Load to Maternal Immune Activation
Maternal immune activation is not caused by epitope counts themselves, but by immune activation events—including exposure-induced innate immune signaling, cytokine release, and placental response—of which sustained antigen presentation is a contributing factor.
MIA is caused by immune signaling. But antigen load determines the breadth and persistence of antigen presentation, the magnitude of T‑cell engagement, and the likelihood of cytokine release and innate–adaptive crosstalk.
Vaccines are designed to be immunogenic. They frequently include adjuvants or molecular designs that enhance innate immune activation and antigen presentation. Each vaccination event constitutes a discrete immune activation episode. When such episodes occur during pregnancy, they overlap temporally with known windows of fetal vulnerability (Atladóttir, 2010; Al-Haddad, 2019).
The literature on MIA demonstrates that immune activation timing and intensity matter. The literature on vaccine immunology demonstrates that these products reliably activate innate and adaptive immune pathways. What has not been adequately studied is the intersection: direct measurement of MIA biomarkers following maternal vaccination and their relationship to fetal outcomes.
Resolving the intersection between maternal vaccination and MIA requires direct measurement rather than inference. Priority research directions include trimester-resolved cytokine profiling following vaccination, placental signaling biomarkers such as STAT3 activation, comparative immune signatures between vaccination and infection during pregnancy, and longitudinal follow-up of offspring neurodevelopmental outcomes.
Conclusion
Maternal immune activation during pregnancy is a documented developmental risk factor. Serious infections during pregnancy are relatively uncommon. In contrast, current CDC recommendations explicitly schedule hundreds to thousands of multiple immune activation events during pregnancy through vaccination.
Claims that these recommendations involve “very few antigens” are based on a categorical error: counting pathogens rather than proteins and ignoring peptide‑level immune recognition. When antigen biology is correctly accounted for, the CDC pregnancy vaccination schedule involves exposure to 15–17 protein antigens comprising on the order of twenty thousand peptide windows, from which dozens of immunogenic epitopes per antigen are realized in human populations.
This does not establish harm. It establishes that immune activation during pregnancy is neither minimal nor negligible and therefore warrants direct study. Assertions of safety that do not measure maternal immune activation itself are incomplete. The burden of proof lies with those who recommend immune activation during pregnancy to demonstrate, rather than assume, that it does not meaningfully intersect with known MIA pathways.
Immune activation during pregnancy is a documented developmental modifier. Vaccination during pregnancy is a designed immune activation event. When these two facts coexist, the appropriate scientific response is measurement, not assumption. Claims of safety that do not include direct assessment of maternal immune activation—and its placental and fetal correlates—do not meet the standard of evidence-based medicine. The burden of proof lies with those asserting immunologic insignificance to demonstrate it empirically.
Or they must accept that others will now do the research they never wanted.
That is what evidence‑based medicine requires.
Popular Rationalists: Starting in early 2026 (Time/Day TBD) Dr. Xavier Figueroa and Tori Alexander will be hosting a session of my course “Autoimmunity and Human Health”
Citations
Smith SE, Li J, Garbett K, Mirnics K, Patterson PH. Maternal immune activation alters fetal brain development through interleukin-6. J Neurosci. 2007;27(40):10695–10702.
Hsiao EY, Patterson PH. Activation of the maternal immune system induces endocrine changes in the placenta via IL-6. Brain Behav Immun. 2011;25(4):604–615.
Wu WL, Hsiao EY, Yan Z, Mazmanian SK, Patterson PH. The placental interleukin-6 signaling controls fetal brain development and behavior. Brain Behav Immun. 2017;62:11–23.
Tioleco N, et al. Maternal Infection and Risk of Autism Spectrum Disorder: A Meta-Analysis. Autism Res. 2021;14(6):1296–1316.
Antoun M, et al. Fever during pregnancy as a risk factor for neurodevelopmental disorders: results from a systematic review and meta-analysis. Mol Autism. 2021;12(1):60.
Al-Haddad BJS, et al. Long-term Risk of Neuropsychiatric Disease After Exposure to Infection In Utero. JAMA Psychiatry. 2019;76(6):594–602.
Atladóttir HÓ, Thorsen P, Østergaard L, et al. Maternal infection requiring hospitalization during pregnancy and autism spectrum disorders. J Autism Dev Disord. 2010;40(12):1423–1430.
Brown AS, Sourander A, Hinkka-Yli-Salomäki S, et al. Elevated maternal C-reactive protein and autism in a national birth cohort. Mol Psychiatry. 2014;19(2):259–264.
Canetta S, Sourander A, Surcel HM, et al. Elevated maternal C-reactive protein and increased risk of schizophrenia in a national birth cohort. Am J Psychiatry. 2014;171(9):960–968.
Zhou Y-Y, et al. Maternal infection during pregnancy and the risk of psychosis in the offspring: a meta-analysis. J Psychiatr Res. 2021;135:28–36.
Saatci D, et al. Maternal infection in gestation increases the risk of non-affective psychosis in offspring: a meta-analysis. J Psychiatr Res. 2021;139:125–131.
Cassotta A, et al. Deciphering and predicting CD4+ T cell immunodominance of influenza virus hemagglutinin. J Exp Med. 2020;217(10):e20200206.
Mateus J, Grifoni A, Tarke A, et al. Selective and cross-reactive SARS-CoV-2 T cell epitopes in unexposed humans. Science. 2020;370(6512):89–94.
Parker R, et al. Mapping the SARS-CoV-2 spike glycoprotein-derived peptidome presented by HLA class II on dendritic cells. Cell Rep. 2021;35(8):109179.
van Bleek GM, Blais S, Sette A. Examining the human CD4+ T cell response to the respiratory syncytial virus fusion protein: identification of epitopes and characterization of their HLA restriction. J Immunol. 2004;172(6):4090–4097. https://pubmed.ncbi.nlm.nih.gov/12502814/
da Silva Antunes R, et al. Definition of Human Epitopes Recognized in Tetanus Toxoid and Development of an Assay Strategy to Detect Ex Vivo Tetanus CD4+ T Cell Responses. PLoS One. 2017;12(1):e0169086.
Babon JA, Cruz J, Orphin L, et al. Genome-wide screening of CD4 T cell responses to influenza virus in humans reveals pervasive patterns of recognition. J Immunol. 2012;188(10):5374–5385.

The vaccine industry's "Just a few antigens so no harm done by vaccinations during pregnancy" position reminds me of Paul Offit's "A baby's immune system can respond to 10,000 antigens at once, so there's no problem with giving a baby several different vaccinations at the same time."
The vaccine industry's crude "Vaccines are safe, effective, and necessary" magic tricks depend on the public's ignorance, gullibility, and "desire to believe."
The COVID spikejab's widespread horrific adverse effects are opening many people's minds to the possibility that vaccines aren't the innocent miracle workers they are promoted as. So we have an opportunity to educate people. Can articles like this one be boiled down to a paragraph or two, or better yet, a short "cartoon strip" with simple eye-catching illustrations, and launched into social media? Maybe "corrective knowledge" about vaccines could go viral.
On a related topic, remember the Zika virus that was blamed for causing microcephaly in Brazilian babies even though it had been around for decades and never done that before? An odd thing was that this epidemic of microcephaly just happened to occur right after a big push to give pregnant women in that region the tetanus/diphtheria/pertussis vaccine. Hmmm...
And then some anti-vaxxer in the U.S. looked up statistics on microcephaly in the U.S. and -- goodness! -- discovered that the incidence of microcephaly in U.S. babies increased by a factor of six after tetanus/diphtheria/pertussis vaccination was recommended for pregnant women in the U.S.
Oddly, not long after that alarming information was put out in social media, the statistics on microcephaly in U.S. babies *prior* to this vaccination recommendation were "disappeared" from the website where this anti-vaxxer had seen them.
Just another magic trick by the vaccine industry. But the "microcephaly effect" should certainly be included when we talk about risks of vaccination during pregnancy, don’t you think?
Extremely important topic. Thank you. This is a drum that needs to be beat again and again. No vaccines during pregnancy to avoid increasing what we don't want: immune activation. The burden of proof is on those who want to vaccinate pregnant women and they have come no where near "clearing the bar". Again, thank you for your clarity and precision.